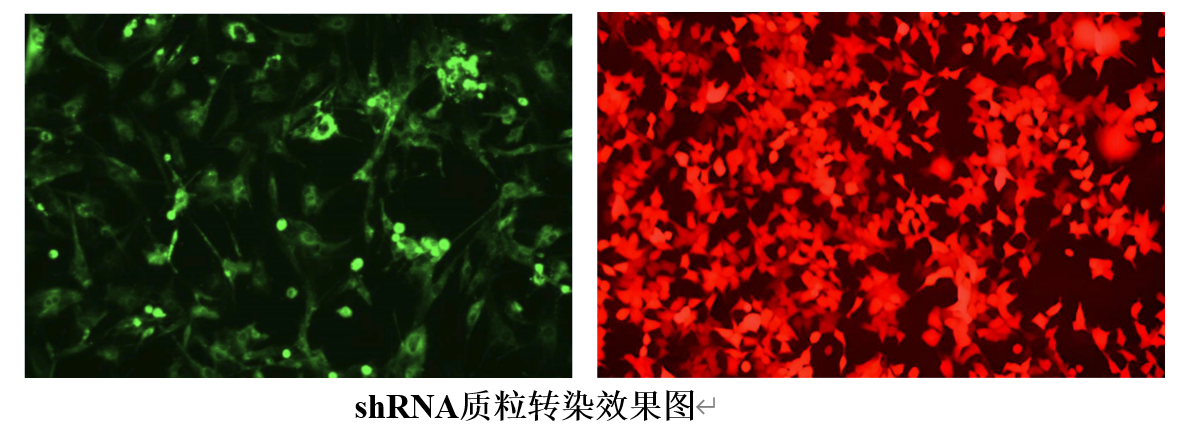
0572-2022975




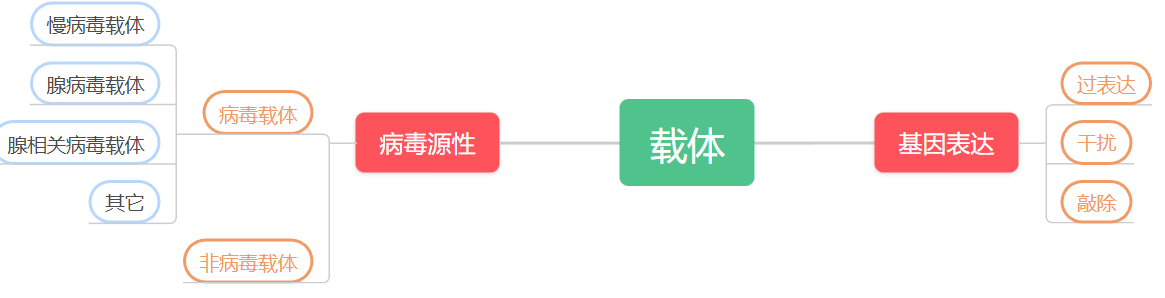
质粒(Plasmid),染色体外能进行自主复制的遗传单位(脱氧核糖核酸,DNA),是目前基因功能研究中常用的一种研究工具,根据其对基因表达的用途可分为过表达质粒、干扰质粒和敲除质粒;根据其病毒源性,又可分为病毒和非病毒质粒(如下图)。
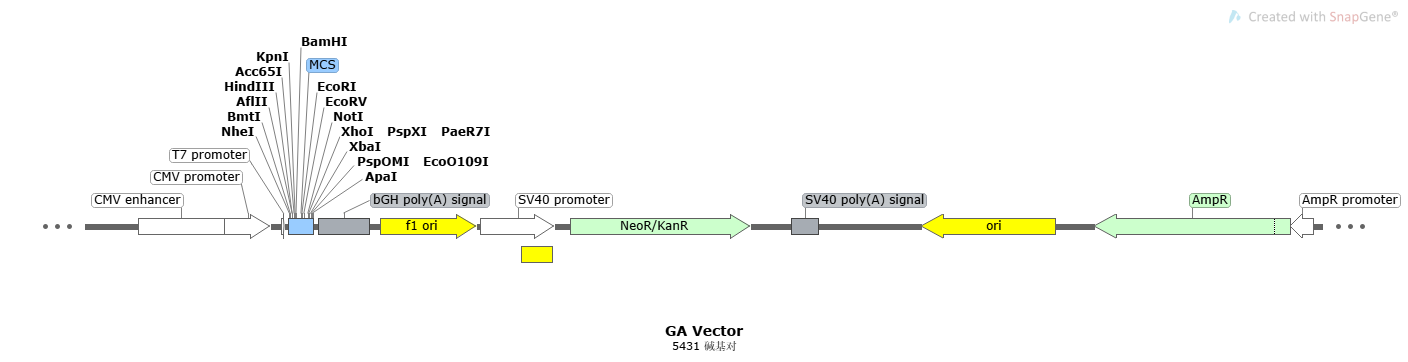
常规质粒骨架:
复制起点(Ori):在质粒骨架中常以Ori结尾标记,复制方向为Ori箭头的方向。分为原核Ori和真核Ori,决定了质粒的宿主及质粒的拷贝数,具有相同Ori的质粒不可共转染(质粒的不相容性)。Ori处的A和T特别多,相对氢键少,不稳定,DNA容易解旋,因此就容易和引物结合,成为转录的起点或复制的起点。|
组织 |
启动子 |
长度 |
备注 |
|
神经 |
NSE |
|
神经元细胞特异启动子 |
|
mDlx |
|
抑制性的中间神经元启动子 |
|
|
ChAT |
|
胆碱能神经元的特异启动子 |
|
|
hSyn |
471 bp |
神经元特异性启动子hSyn。 |
|
|
mecp2 |
|
短的神经元特异性启动子。 |
|
|
c-fos |
|
兴奋神经元特异性启动子。 |
|
|
CaMKIIa |
1.2 kb |
兴奋性谷氨酸能神经元启动子 |
|
|
hVGAT |
|
GABA能神经元/中间神经元特异性启动子 |
|
|
gfaABC1D |
|
星形胶质细胞特异启动子 |
|
|
lba1 |
|
小胶质细胞特异启动子 |
|
|
NSE |
|
神经原特异启动子 |
|
|
GFAP104 |
|
星形胶质细胞特异启动子 |
|
|
AlDH1L1 |
|
丘脑中星形胶质细胞特异启动子 |
|
|
MBP |
|
髓鞘碱性蛋白启动子,表达在少突胶质细胞 |
|
|
Somatostat |
1.2 kb |
GABA能神经元特异性启动子 |
|
|
CNP |
|
少突胶质细胞或施万细胞特异性表达启动子 |
|
|
心脏 |
cTNT |
702 bp |
心脏特异性启动子 |
|
aMHC |
400 bp |
小鼠肌球蛋白重链启动子 |
|
|
SM22a |
441 bp |
平滑肌细胞特异性启动子 |
|
|
MCK |
|
肌酸激酶启动子 |
|
|
Nkx2.5 |
|
心肌细胞早期特异启动子 |
|
|
肌肉 |
MCK |
1.3 kb |
小鼠肌肉细胞特异性启动子 |
|
MHCK7 |
|
肌肉细胞特异性启动子 |
|
|
MyOG |
|
成肌细胞特异性启动子 |
|
|
内皮 |
TIE1 |
|
内皮细胞特异启动子 |
|
血管内皮 |
EnSm22a |
|
血管平滑肌特异启动子 |
|
ICAM2 |
140 bp |
血管内皮特异启动子 |
|
|
单核巨噬细胞 |
CD68 |
|
单核巨噬细胞特异启动子 |
|
巨噬细胞 |
F4/80 |
|
巨噬细胞特异启动子 |
|
肝脏 |
TBG |
460 bp |
肝脏特异启动子 |
|
ALB |
2.4 kb |
肝脏特异启动子 |
|
|
ApoEHCR-hAAT |
|
肝脏特异启动子 |
|
|
胰腺 |
ins2 |
|
胰腺β细胞特异启动子 |
|
PDX1 |
|
胰腺β细胞特异启动子 |
|
|
骨骼肌 |
COL2A1 |
|
软骨细胞特异启动子 |
|
mRUNX2 |
|
成骨细胞早期特异启动子 |
|
|
眼睛 |
Rpe65 |
700 bp |
视网膜色素上皮细胞特异启动子 |
|
hVMD2 |
642 bp |
视网膜色素上皮细胞特异启动子 |
|
|
肾脏 |
NPHS1 |
|
肾脏特异启动子 |
|
肺 |
SPB |
|
肺上皮细胞特异启动子 |
|
SPC |
|
肺上皮细胞特异启动子 |
|
|
角质层 |
K14 |
|
角化细胞特异启动子 |
|
脂肪 |
KAPB4 |
|
脂肪细胞特异启动子 |
|
广谱启 |
CAG |
|
强启动子,常用于体内表达 |
|
EFES |
|
EF1A的短版本 |
|
|
EF1a |
1.2 kb |
效果弱于CMV,但体内实验效果好 |
|
|
CMV |
600 bp |
最常用的强启动子 |
|
|
CBA |
|
类似于CAG,是人工启动子 |
|
|
PGK |
|
效果弱于CMV,体内效果好 |
|
|
诱导型 |
TRE |
|
四环素抗性诱导启动子 |
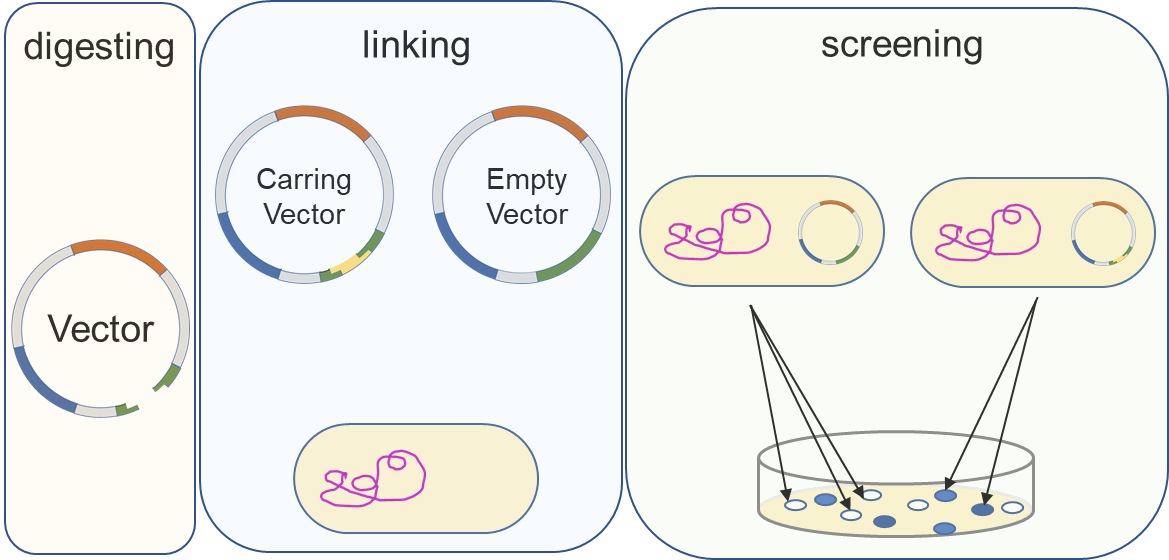
引物设计:通过NCBI(https://www.ncbi.nlm.nih.gov/)的Gene模块获得基因序列,NCBI的blast模块设计对应的引物。引物设计注意事项:GC最好控制在40~60%;长度最好18~30 nt;内部重复及互补结构最好不大于3;Tm最好控制在55~60℃,两天引物Tm值差异最好不大于3;3’端结尾尽可能是G/C,避免为A;引物5’端可添加酶切位点或其它序列,如添加酶切位点,5’端需添加保护碱基(一般4个),注意避免酶切位点原核甲基化,影响后续酶切实验。
PCR扩增(以PrimeSTAR® Max DNA Polymerase为例):|
试 剂 |
使 用 量 |
终 浓 度 |
|
PrimeSTAR Max Premix(2X) |
25 μl |
1X |
|
Primer 1 |
10 ~ 15 pmol |
0.2~0.3 μM |
|
Primer 2 |
10 ~ 15 pmol |
0.2~0.3 μM |
|
Template |
<200 ng* |
|
|
灭菌水 |
Up to 50 μl |
|
|
|
温度 |
时间 |
循环 |
|
预变性 |
98℃ |
30 sec |
1 |
|
变性 |
98℃ |
10 sec |
30~35 |
|
退火 |
60℃ |
5-15 sec |
|
|
延伸 |
72℃ |
30~60 sec/kb |
|
|
延伸 |
72℃ |
5 min |
1 |
|
保存 |
4℃ |
~ |
- |
|
ddH2O |
16-x μL |
|
10×缓冲液 |
2 μL |
|
DNA |
x μL |
|
酶 1 |
1 μL |
|
酶 2 |
1 μL |
|
Total |
20μL |
|
|
1× |
0.5× |
|
线性化质粒DNA |
20-100 ng |
50 ng |
|
Insert DNA |
1:1 to 5:1 |
3:1 |
|
Buffer |
2 µL |
1 µL |
|
T4 DNA Ligase |
1/5 Weiss U |
0.2/1 µL |
|
ddH2O |
to 10 µL |
|
|
Total |
20μL |
10 μL |
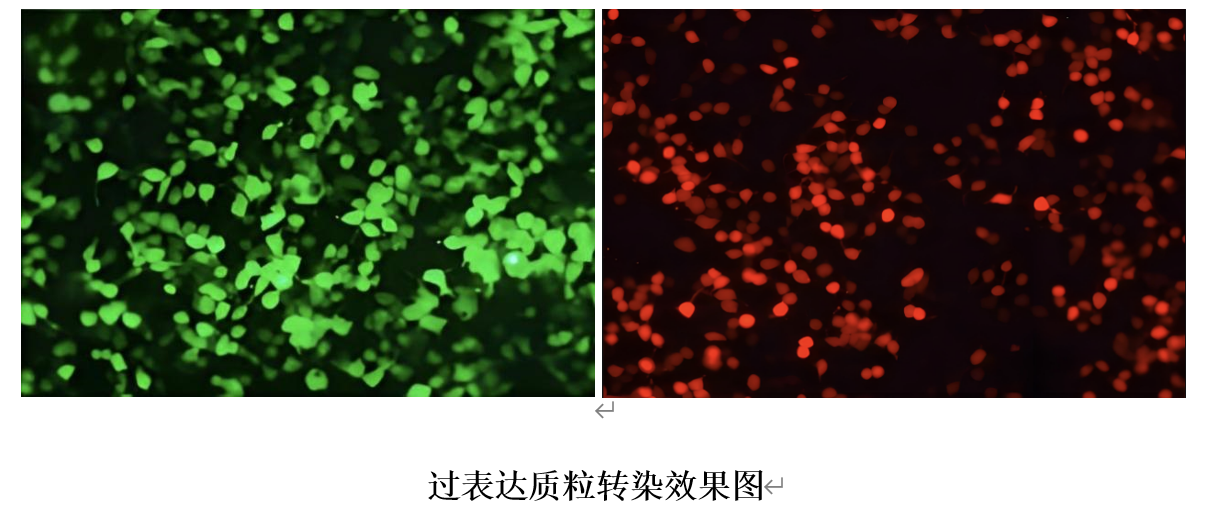
|
产品类型 |
产品组分及要求 |
|
单条shRNA |
单条shRNA |
|
shRNA三保一 |
三条目的基因shRNA,空载对照 |
|
shRNA三保一升级版 |
三条目的基因shRNA,空载对照,转染试剂 |
|
shRNA四保一 |
四条目的基因shRNA,空载对照 |
|
基因过表达质粒 |
根据不同实验目的进行改造 |
|
质粒抽提 |
去内毒素 |
|
plko.1-PURO |
无荧光 |
PURO抗性 |
|
plko.1-NEO |
无荧光 |
NEO抗性 |
|
plko.1-Hygro |
无荧光 |
Hygro抗性 |
|
plko.1-copGFP-PURO |
绿色荧光 |
PURO抗性 |
|
plko.1-mCherry-PURO |
红色荧光 |
PURO抗性 |
|
tet-plko-PURO |
tet诱导 |
PURO抗性 |
|
plko.1-CMV-copGFP-PURO |
绿色荧光 |
PURO抗性 |